Mouse ON-TARGETplus siRNA Library - Druggable Genome
siRNA designed and modified for greater specificity
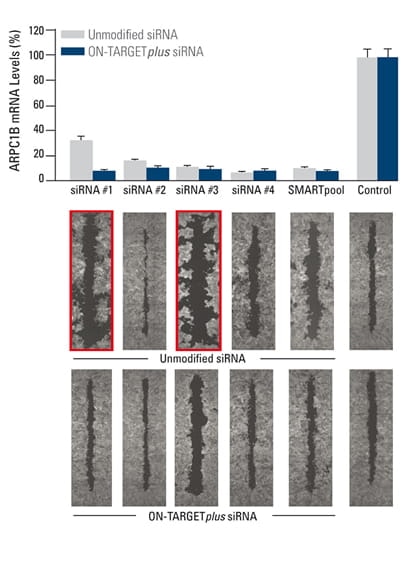
An arrayed collection of siRNA reagents for RNAi screening, targeting mouse genes that are potential therapeutic targets. ON-TARGETplus siRNA is guaranteed to silence and is modified to reduce off-targets for fewer false positives.
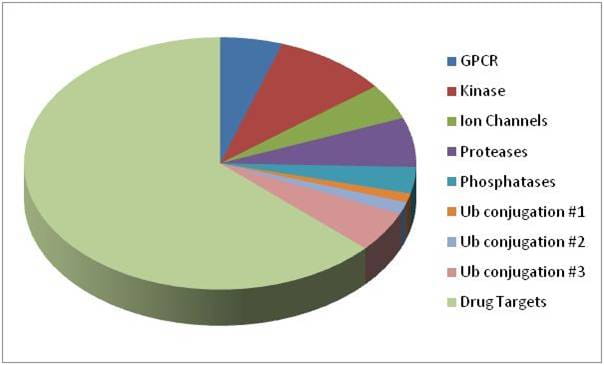
The ON-TARGETplus Druggable Genome library contains the genes that are considered potential targets for therapeutics, including kinases, proteases, phosphatases, GPCRs, ion channels, ubiquitin ligases, and genes involved in senescence, autophagy, DNA repair, and characterized nuclear receptors. This extensive library is delivered as nine distinct subset siRNA libraries in the 96-well format (see Supporting Data tab). In the 384-well format, the first eight subsets are plated continuously to reduce partial plates.
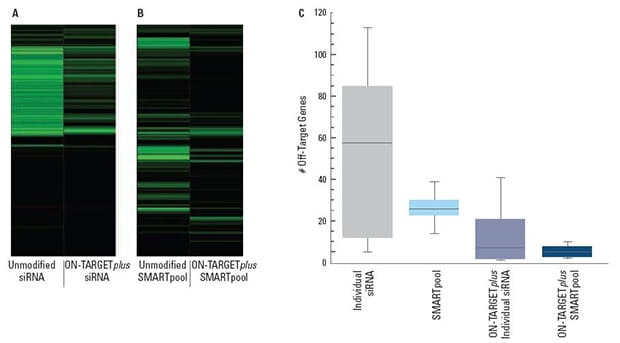
The ON-TARGETplus siRNA designs and modifications reduce off-target effects while maintaining high-silencing potency for high-confidence screening results.
Highlights
- Patented dual-strand ON-TARGETplus modification pattern on all siRNAs to reduce off-targets
- Sense strand is modified to prevent interaction with RISC and favor antisense strand uptake
- Antisense strand seed region is modified to destabilize off-target activity and enhance target specificity
- Available as SMARTpool siRNA reagents or a Set of 4 siRNAs in 96 or 384-well plates
- Guaranteed target gene knockdown (see Specifications tab)
Nine subsets make up the mouse druggable genome siRNA library
|
|