Our premium shRNA products use a microRNA-adapted shRNA design to promote more efficient cellular processing and reduce toxicity during RNAi experiments. Visit our shRNA applications page to learn more.
However, efficient gene silencing depends on both the shRNA design and expression level. Therefore, promoter selection is also critical to drive sufficient shRNA expression in your specific cell line.
SMARTvector™ – shRNA tailored to your experiment
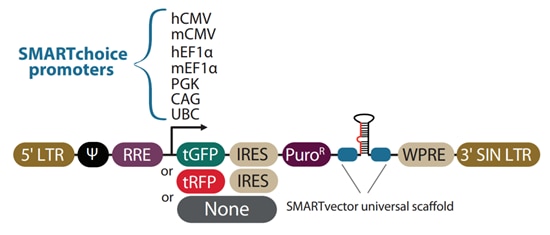
The SMARTvector backbone allows you to choose from seven promoters and three reporter options to optimize shRNA for your specific cells and experimental requirements.
Designed using our most advanced microRNA-based shRNA-specific rational design algorithm, SMARTvector lentiviral shRNA expressed under an optimized promoter produces highly potent and specific silencing. Identify the optimal promoter for your cell line with the SMARTchoice promoter selection plate.
SMARTvector shRNA are also available with a Tet-on® 3G inducible promoter for reversible, controlled gene silencing, which is especially useful when studying the function of essential genes.
GIPZ/TRIPZ collections
GIPZ shRNA and TRIPZ inducible shRNA offer specific gene silencing with minimized cellular toxicity, but lack the SMARTvector's promoter/reporter flexibility. Both GIPZ and TRIPZ collections were developed in collaboration with Gregory Hannon (Cold Spring Harbor Laboratories) and Steve Elledge (Harvard).
Order shRNA reagents
SMARTvector lentiviral shRNA
Guaranteed gene silencing with multiple promoter and reporter options.
SMARTvector inducible lentiviral shRNA
Smartvector lentiviral shRNA with Tet-inducible technology for unprecedented silencing control.
GIPZ lentiviral shRNA
Efficient gene silencing with a microRNA-adapted shRNA design.
TRIPZ inducible lentiviral shRNA
Inducible shRNA expression for tightly regulatable RNAi experiments.
TRC lentiviral shRNA
Lentiviral shRNA collection from The RNAi Consortium (TRC) for human and mouse cells.
Helpful resources
Watch video now
Tips for determining functional titer - Measuring functional titer can help you make the most out of your precious lentiviral reagents
Which shRNA reagent is best for you?
Use the table below to assist you in determining the right shRNA reagents for your experimental needs.
| SMARTvector shRNA | GIPZ | TRIPZ | TRC | |
|---|---|---|---|---|
| Species | Human, Mouse, Rat | Human, Mouse | Human | Human, Mouse |
| Promoter | Choice of 7 constitutive* and 4 inducible promoters | Human CMV Pol II | TRE-min-CMV | U6 Pol III |
| Vector backbone | Lentiviral | Lentiviral | Lentiviral | Lentiviral |
| Stem-loop format | MicroRNA-adapted | MicroRNA-adapted | MicroRNA-adapted | Simple |
| Fluorescent reporter gene | GFP/RFP | GFP | RFP | |
| Guaranteed silencing** | ||||
| In vivo RNAi | ||||
| Create stable cell lines | ||||
| Recommended for primary and non-dividing cells | ||||
| Inducible expression | ||||
| Whole genome library availability | Human, Mouse, Rat | Human, Mouse | Human | Human, Mouse |
| Formats | Bacterial glycerol stock, High-titer lentiviral particles, Gene families and pathways | Bacterial glycerol stock, Arrayed library format, Gene families and pathways, High-titer lentiviral particles, shRNA Starter Kit | Bacterial glycerol stock, Arrayed library format, Gene families and pathways, shRNA Starter Kit | Bacterial glycerol stock, Arrayed library format, Gene families and pathways |
*Some promoter options may only be available as custom products or upon request.
**For SMARTvector, GIPZ, and TRIPZ lentiviral shRNAs, at least one out of three constructs is guaranteed to reduce target mRNA levels by 70% or more when used in combination with the appropriately matched non-targeting and positive controls.


SMARTvector shRNA reagents & controls
Critical to any gene silencing experiment, shRNA controls enable accurate interpretation for reliable, reproducible results.
-
SMARTvector lentiviral shRNA
Guaranteed gene silencing with multiple promoter and reporter options.
-
SMARTvector lentiviral controls
Positive and negative constitutive shRNA controls with choice of seven promoters and three reporter options.
-
SMARTvector inducible lentiviral shRNA
Smartvector lentiviral shRNA with Tet-inducible technology for unprecedented silencing control.
-
SMARTvector inducible lentiviral controls
Inducible expression of positive and negative controls with your choice of four promoters and two reporters.
-
SMARTchoice promoter selection plate
Identify the optimal SMARTvector promoter option for your cell line.
shRNA collections & controls
-
GIPZ lentiviral shRNA
Efficient gene silencing with a microRNA-adapted shRNA design.
-
GIPZ lentiviral controls
Validated collection of GIPZ positive and negative controls for a well-designed RNAi experiment.
-
TRIPZ inducible lentiviral shRNA
Inducible shRNA expression for tightly regulatable RNAi experiments.
-
TRIPZ inducible lentiviral controls
Positive and negative TRIPZ Inducible Lentiviral shRNA Controls for setting the experimental window.
-
TRC lentiviral shRNA
Lentiviral shRNA collection from The RNAi Consortium (TRC) for human and mouse cells.
-
TRC lentiviral controls
Positive shRNA control targeting GFP and empty lentiviral vector negative control available as glycerol stocks
Pooled shRNA screening libraries
A pooled lentiviral screen can be performed to identify genes that regulate cellular responses and signaling pathways, or to discover novel gene functions. Pooled screening libraries can consist of as few as 50 constructs up to many thousands. In contrast to the costly automated techniques that are required to screen using individually arrayed reagents, pooled screening libraries allow the researcher to transduce and screen a population of cells within a few tissue culture dishes.
-
Custom pooled lentiviral screening libraries
Flexible collections of lentiviral shRNA for customizable RNAi knockdown screens.
-
SMARTvector lentiviral shRNA pooled libraries
High-titer pooled screening libraries of constitutive SMARTvector Lentiviral shRNAs for pre-defined gene libraries in human, mouse, and rat. Choose from seven promoters and three reporter options to select the best format for your experiment.
-
SMARTvector inducible lentiviral shRNA pooled libraries
High-titer pooled screening libraries of SMARTvector Inducible Lentiviral shRNAs for pre-defined gene libraries in human, mouse, and rat. Choose from four promoters and two reporter options to select the best format for your experiment.
Arrayed shRNA screening libraries
Delivered as glycerol stocks in 96-well format; these libraries are ideal for arrayed screening formats to elucidate gene activity and study specific pathways or gene families.
-
Custom cherry-pick clone libraries
Upload your gene list to build your own custom arrayed shRNA library.
-
GIPZ lentiviral shRNA libraries
Genome-wide collections for high-throughput shRNA screening
-
TRIPZ inducible lentiviral shRNA libraries
Genome-scale TRIPZ Inducible Lentiviral shRNA library arrayed in 96-well plates
-
TRC lentiviral shRNA libraries
Simple hairpin shRNAs in the pLKO.1 lentiviral vector designed by The RNAi Consortium (TRC)
The SMARTvector functional guarantee
When you purchase a minimum of three SMARTvector lentiviral shRNAs to the same protein-coding gene target using the optimal SMARTchoice promoter for your cell type, at least one of the shRNA constructs will reduce target mRNA levels by 70% or more when used with the vector matched non-targeting control and GAPD or PPIB positive control. Optimal promoters should be determined with the SMARTchoice promoter selection plate. mRNA levels should be measured with RT-qPCR or similar quantitative mRNA analysis no earlier than 72 hours post-transduction.
The guarantee only applies to catalog numbers V3S*xxxx and does not apply to any SMARTvector shRNAs targeting lncRNA transcripts.
GIPZ/TRIPZ functional guarantee
When you purchase a minimum of three shRNA to the same target, at least one of the shRNA constructs will reduce target mRNA levels by 70% or more when used with Starter Kit protocols and normalized for delivery efficiency. Delivery conditions should be confirmed using appropriate positive controls provided in the kit and the percent-knockdown should be compared to cells using the negative control (non-targeting) shRNA.
Please note that the GIPZ Lentiviral shRNA and TRIPZ Inducible Lentiviral shRNA are not compatible with third generation packaging systems, such as ViraPower from Invitrogen. We recommend the Trans-Lentiviral Packaging System to general lentiviral particles for transduction of GIPZ and TRIPZ shRNA.