Mouse siGENOME siRNA Library - Proteases
Trusted for superior silencing since 2002
An arrayed siRNA collection targeting mouse proteases. siGENOME siRNA is a cost-effective choice for RNAi screening. Available as SMARTpool or 4 individual siRNA reagents.
With the Mouse siGENOME Proteases siRNA library, researchers receive siRNAs targeting enzymes with known or predicted protease activity. Proteases, also known as peptidases or proteinases, constitute enzymes whose role is to hydrolyze protein peptide bonds, and through this mechanism, they mediate a great number of physiological processes, including embryonic development, wound healing, the immune response, apoptosis, hormonal control, and many others.
siGENOME siRNA reagents are globally recognized and trusted for highly effective performance in RNAi screening applications. Our expertise in siRNA design, experimental optimization, and siRNA screening strategies provide end-to-end support of your screening efforts.
Highlights
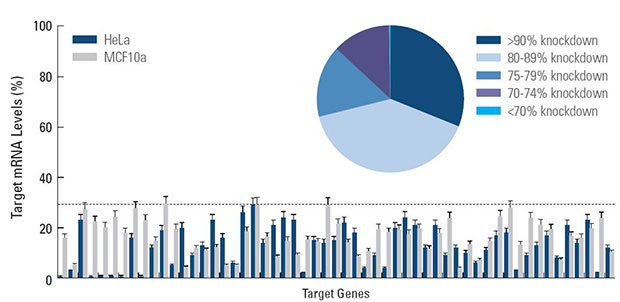
- Guaranteed target knockdown (see Guarantee tab)
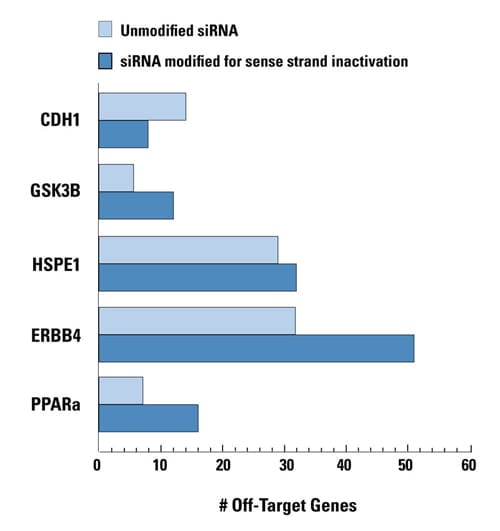
- Antisense strand loading into RISC ensured by thermodynamic analysis and selective application of a sense strand-blocking modification (ON-TARGET)
- Available as SMARTpool or Set of 4 siRNA reagents arrayed in 96-well plates
Gene Targets
For a complete list of target genes in this siRNA Library, please contact Scientific Support.