Mouse siGENOME siRNA Library - Protein Kinases
Trusted for superior silencing since 2002
An arrayed siRNA collection targeting mouse kinases. siGENOME siRNA is a cost-effective choice for RNAi screening. Available as SMARTpool or 4 individual siRNA reagents.
The Mouse siGENOME Protein Kinase siRNA Library includes siRNA reagents directed against phylogenetically-related kinases. Protein kinases are key regulators of cell function and are known to direct the activity, localization, and function of many proteins. They are prominent in signal transduction and are crucial for the coordination of complex cellular functions such as cell cycle regulation, gene transcription, and apoptosis.
siGENOME siRNA reagents are globally recognized and trusted for highly effective performance in RNAi screening applications. Our expertise in siRNA design, experimental optimization, and siRNA screening strategies provide end-to-end support of your screening efforts.
Highlights
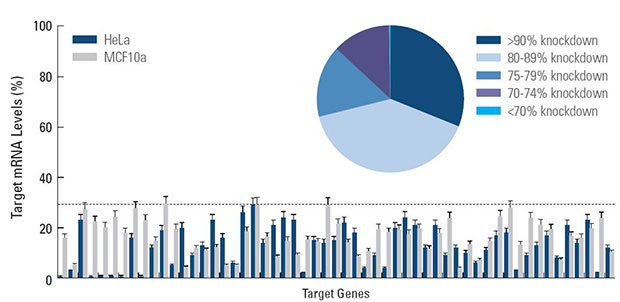
- Guaranteed target knockdown (see Specifications tab)
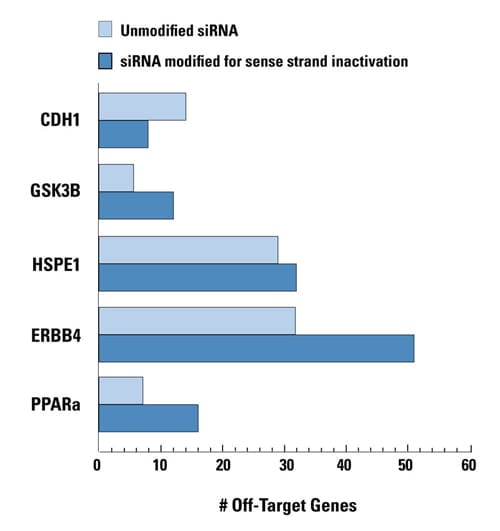
- Antisense strand loading into RISC ensured by thermodynamic analysis and selective application of a sense strand-blocking modification (ON-TARGET)
- Available as SMARTpool or Set of 4 siRNA reagents arrayed in 96-well plates
Gene Targets
For a complete list of target genes in this siRNA Library, please contact Scientific Support or your local Sales Representative.