Mouse siGENOME siRNA Library - Ubiquitin Conjugation Subset 3
Trusted for superior silencing since 2002
An arrayed siRNA collection targeting mouse RING-finger E3 Ligases. siGENOME siRNA is a cost-effective choice for RNAi screening. Available as SMARTpool or 4 individual siRNA reagents.
The Mouse siGENOME Ubiquitin Conjugation 3 siRNA library targets RING finger and RING finger-like E3 ligases. These E3s form a site of interaction with E2s through a platform generated as a consequence of coordination of two zinc ions. U-box E3s conform like RING-finger proteins except that they do so through salt bridges and other interactions that do not involve coordination of zinc. This set contains A20-like proteins, which contain a single coordination site for zinc, and siRNA for well-known E3s,such as BRCA1, MDM2, and CBL family members.
siGENOME siRNA reagents are globally recognized and trusted for highly effective performance in RNAi screening applications. Our expertise in siRNA design, experimental optimization, and siRNA screening strategies provide end-to-end support of your screening efforts.
Highlights
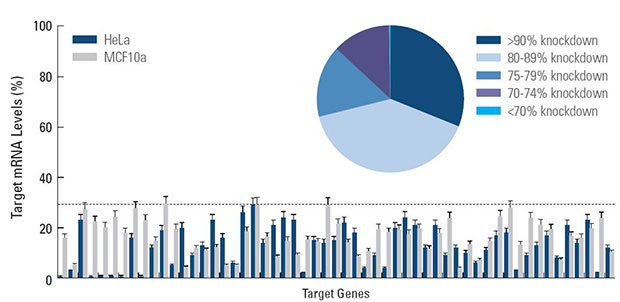
- Guaranteed target knockdown (see Specifications tab)
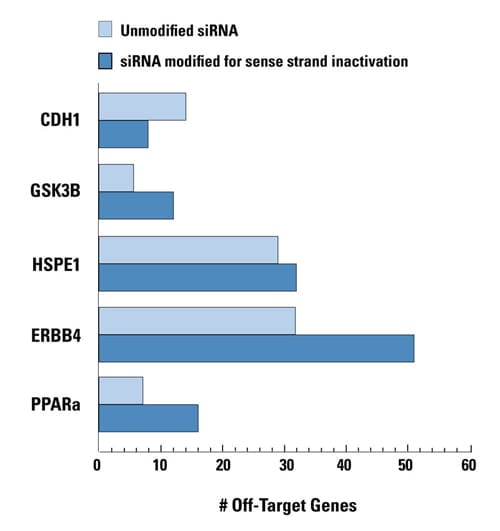
- Antisense strand loading into RISC ensured by thermodynamic analysis and selective application of a sense strand-blocking modification (ON-TARGET)
- Available as SMARTpool or Set of 4 siRNA reagents arrayed in 96-well plates
Gene Targets
For a complete list of target genes in this siRNA Library, please contact Scientific Support or your local Sales Representative.
For a thorough investigation of the ubiquitin pathway, you may also consider these additional libraries: