Strict-R Inducible CRISPRa Lentiviral System
How it works
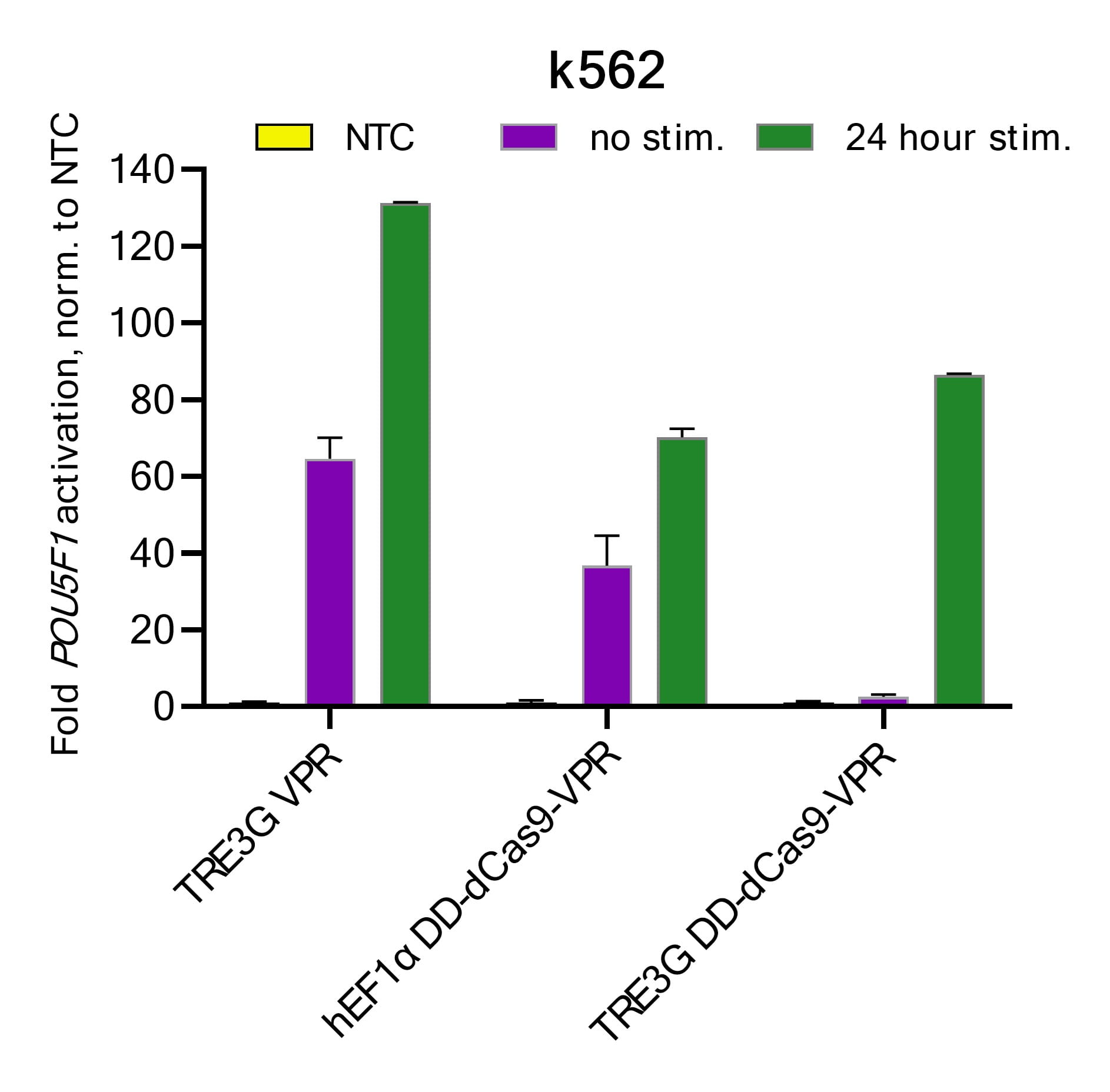
System OFF: In the absence of doxycycline and Shield1, the system remains inactive. Any residual transcription from the TRE3G promoter produces degron-tagged dCas9-VPR, which undergoes rapid proteasomal degradation, minimizing unintended background activation.
System ON: Doxycycline administration activates the Tet-On 3G transactivator, inducing robust dCas9-VPR expression. Shield1 addition stabilizes the expressed dCas9-VPR protein, allowing accumulation and precise gene activation.
Schematic map of the Strict-R Inducible Lentiviral CRISPRa vector

Transcriptional and post-translational control with the Strict-R Inducible Lentiviral CRISPRa System

Diagram of the Strict-R Inducible Lentiviral CRISPRa System. In the absence of Doxycycline and Shield1, the system is “OFF”. Leaky bursts of transcription from the TRE3G promoter result in the translation of dCas9-VPR fused to a FKBP12-derived destabilizing domain (degron) that tags the protein for rapid proteasomal degradation . The addition of doxycycline induces potent transcription from the TRE3G promoter and the addition of Shield1 stabilizes dCas9-VPR thereby enabling robust target gene activation in the presence of a gene-specific sgRNA. Diagram created with BioRender.com.
Highlights
- Achieve reversible, small molecule-induced activation of target genes, providing tight regulation of gene expression.
- The dCas9-VPR fusion protein maximizes transcriptional activation with minimal off-target effects.
- The integration of the Tet degron system allows for rapid induction of dCas9-VPR expression upon the addition of doxycycline and Shield1, facilitating dynamic, time-controlled experiments.
- This system is expressed from a single lentiviral vector facilitating integration into your experimental workflow.
- Enables wide range of research applications including gene expression studies, functional genomics, and potential therapeutic applications.
- High quality, purified lentiviral particles for direct transduction with minimal cytotoxicity; delivered at titers of ≥ 1 x 107 TU/mL
- Options for either Blastacidin selection or EGFP fluorescent reporter.